ノーウォークウイルス(ノロウイルス)の遺伝子型(2015年改訂版)
(掲載日 2015/9/8)
はじめに
ノーウォークウイルス(ノロウイルス、NoV)に限らず、すべてのウイルスは、電子顕微鏡で可視化しても、クライオ電子顕微鏡観察により3Dイメージを再構築しても、ウイルス粒子自身に血清型番号や、遺伝子型番号などが付いているわけではない。血清型や遺伝子型は、我々人類が何らかの目的を持って分別した結果なのである。
2011年以降、ノロウイルスサイエンティフィックコミッティー(NoV S.C.)により、NoVの流行のメカニズムをより深く研究する目的で、2000年以降、2006/07シーズン、2012/13シーズンに大流行したNoVのGII.4亜株を分別すること、NoVゲノムにコードされる非構造タンパク質部分の特徴を反映させ、遺伝子組換え(genome recombination)にも対応させることを考慮に入れた新規遺伝子型分別法が開発され、2013年にArchives of Virologyに発表された。この遺伝子型分類は、International Norovirus Study Groupに継承され、オランダのMarion Koopmans博士らのNoroNet上のNorovirus Typing Tool(NoV遺伝子型分類ツール)によってWeb上で提供されている。
2015年は、ノロウイルスの主要流行株に大きな動きがあった。2014年12月まで主要流行株を占めていたGII.4が減少に転じ、新たにGII.17 (GII.P17-GII.17、GII.17 Kawasaki variant)が主要流行株に転じたのである1)。主要流行株の入れ替わりは、2006年以降、我々が初めて経験する大きな変化であり2)、2015/16シーズンの立ち上がり、つまり10月下旬~11月終盤にかけてのGII.17の動向には細心の注意を払う必要がある。
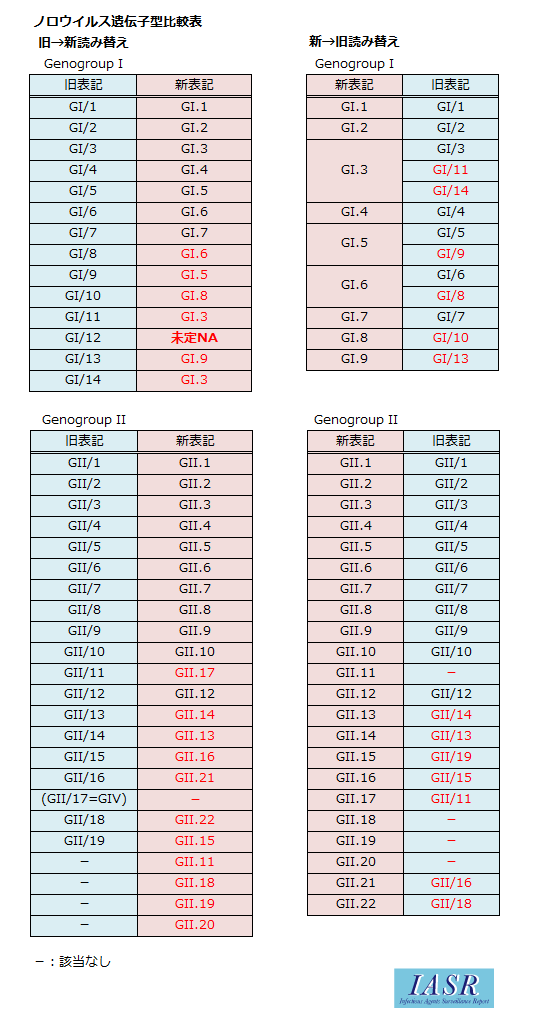
GII.17は、IASRにて用いられてきたスラッシュ区切りの型分類上、GII/11に相当し、GII/17とは異なる遺伝子型である。実は、スラッシュ区切りの型分類上のGII/17は、NoroNet上の新規遺伝子型分別法では、GIV.1に相当するのである。2015年の予想外のGII.17の流行により、スラッシュ区切りの型分類からNoroNet上の新規遺伝子型分別法への変換を、2015/16シーズンより全面的に切り替えることとなった。
本稿ではNoroNet上の新規遺伝子型分別法と、従来我々が提唱し、使用されてきたCapsidNS領域(region D)によるタイピング法3, 4)との比較表をより変換が分かり易く更新するとともに、GII.17のタイピングについて対応したので、案内する (比較表参照)。
{kind=link}
なお、我々が通常使用している“ノロウイルス”という呼び名は、ウイルス属名を示すものであり、ウイルス種名を示すものではない。ノロウイルス属に他のウイルス種が発見されると、この呼び方を改める必要が出てくるが、マウスに感染するノーウォークウイルスをマウスノロウイルス(Murine Norovirus)5)と呼ぶこと、ノーウォークウイルスを表す言葉として、ノロウイルスが多くの学術論文に使用されていること、この慣例に従い、以降本稿ではノロウイルス(NoV)を使用する。
新しい遺伝子型分類法(genotyping)
ウイルスのFamily(科)とGenus(属)は、形態学的特徴、病原性、ゲノムの構造などを加味して、国際ウイルス命名委員会(International Committee on Taxonomy of Viruses; ICTV、http://www.ICTVonline.org/index.asp)が決定する。ノロウイルスは、ICTVのウイルスデータベース; ICTVdBの、カリシウイルスの項目(http://www.ncbi.nlm.nih.gov/ICTVdb/ICTVdB/12000000.htm)に、カリシウイルス科に属する一つのウイルス属として定義されている。カリシウイルス科(Caliciviridae)には、現在5つのウイルス属(genus)の存在が認定されている。ノロウイルス属(Norovirus)、サポウイルス属(Sapovirus)、ベジウイルス属(Vesivirus)、ラゴウイルス属(Lagovirus)、ネボウイルス属(Nebovirus)である。ノロウイルス属(Norovirus)は、ノーウォークウイルス種(Norwalk viruses)を唯一の種(Species)として持つ。ICTVのCaliciviridae Study Group(http://www.ictvonline.org/subcommittee.asp?committee=14)は、Species以下の分別方法には原則として関与しない。NoV S.C.は、ICTVの関与しないSpecies以下の分別方法に関して議論を繰り返し、合意に基づいて“Proposal for a unified norovirus nomenclature and genotyping”6)をまとめた。以下に内容を概説した。
NoVは、約7,500塩基のプラス一本鎖RNAをゲノムに持つウイルスである。NoVのゲノムには3つの蛋白質コード領域(open reading frame; ORF)が存在しており、ORF1;非構造蛋白質、ORF2;構造蛋白質(キャプシド蛋白質;VP1)、ORF3;構造蛋白質(VP2) をコードしている。NoVのゲノム塩基配列は多様性に富んでおり、ゲノム塩基配列の相同性に基づき5種類のgenogroup(GI、II、III、IV、V、これらのうちGVはマウスノロウイルス;MNV)に分類されている。
a)ORF2(キャプシド蛋白質; VP1コード領域)の遺伝子型分類
これまで、5種類のgenogroup以下のクラス、つまりNoVの遺伝子型は、ウイルスの抗原性の違いを推定するために、VP1の完全長塩基配列をpairwise distanceのカットオフ値に基づいて決められていた。それを簡便に再現するため、VP1のN末端領域の約300塩基(Capsid N/S region)を標的にして簡便な遺伝子型分類が行われていた。しかし、解析に利用できるノロウイルスの塩基配列データが増加したため、この分類方法で分別できない遺伝子型が増加しつつあった。
“Proposal for a unified norovirus nomenclature and genotyping”では、pairwise distanceによる分子系統解析を否定するわけではないが、矛盾無く分子遺伝学的にNoVの遺伝子型分類を行うためには、ベイジアン法もしくは最尤法(Maximum-likelihood analysisを用いることが望ましいことを示した。また、Fields Virology 6th editionと同調を図り、ORF2の全長塩基配列を用いたgenotypingによりNoV GIに9つの遺伝子型(GI.1~9)、 GIIには、22種類の遺伝子型(GII.1~22)の存在を明らかにした。さらに、本方法のもう一つの利点は、GII.4などに認められた亜株(variants)の分類を実施できる点にある。Maximum-likelihoodによる分子系統解析では、2006~2007年冬期から2009~2010年冬期の流行の主流を担ってきた78.4%のGII.4 亜株を分類可能であり7)、GII.4 Sydney 2012亜株も例外ではない。さらに、2015年年明けから流行し、GII.4を主要流行株の座から降ろしたGII.P17-GII.17(GII.17 Kawasaki variant)(http://www.niid.go.jp/niid/ja/norovirus-m/norovirus-iasrs/5903-pr4273.html参照)も、同様に分類可能であった。これは、ベイジアン法を用いた場合でも同様であった。
b)ORF1(非構造蛋白質コード領域)のポリメラーゼコード領域の遺伝子型分類
NoVでは、ORF1-2ジャンクション領域を基点とする遺伝子の組換えが頻発する。遺伝子組換えを起こしたキメラウイルスの存在を考慮に入れ、NoVを理解するためには、ORF1-2ジャンクション領域を含むようにORF1からORF2にかけて連続した断片となるように逆転写PCR(RT-PCR)で増幅して分子系統解析する必要がある。
“Proposal for a unified norovirus nomenclature and genotyping”では、ORF1の最下流にコードされているRNA dependent RNA polymerase; RdRp or polymeraseの部分の1,300ntとORF2/VP1領域を対照にし、それぞれの遺伝子型分類を行うことを提案した。同時に両領域の遺伝子型を表現可能な新規命名法を提唱した。この提案は、NoVの分子進化をよりよく理解し、キメラウイルス出現がヒト集団でのNoVの流行にどのように関係するのかを調べるために有用である。また、我々のグループは、NoV非構造タンパク質の一種p22(3A-like protein)の機能解析の結果、p22が細胞の蛋白質の小胞輸送システムを乱し、細胞にダメージを与えことを示した8, 9)。これらの発見もまた、非構造タンパク質領域を標的とした遺伝子型分類がNoVの病原性発現機構の研究に有用であることを示唆している。
RdRp領域を用い、ベイジアン法もしくはMaximum-likelihoodで遺伝子型分類を行うと、166株は、12個のクラスターと12以上の一株のみの遺伝子配列からなる枝に分岐した。これらクラスターの幾つかは、いかなるVP1領域とも関連性の無いクラスターであった。また、異なるVP1領域と繋がるクラスター(たとえばRdRp領域がGII.11のクラスターは、VP1領域がGII.11のクラスターとはもちろんのこと、GII.19のクラスターとも繋がっていた)が存在した。これらは、キメラウイルスの存在を示している。
c) NoroNetのNoV遺伝子型分類ツールを用いた遺伝子型分類
さて、それでは、実際の遺伝子型分類をどのように行うのかを示す。まずは、ORF1の最下流のRdRp領域からORF2のできればストップコドンまでをRT-PCRで増幅する。その後、塩基配列を決定し、NoV遺伝子型分類ツール(http://www.rivm.nl/mpf/norovirus/typingtool)にインターネットでアクセスする。ガイドに従い、配列をコピー&ペーストして、解析開始ボタンを押すと、RdRp領域、VP1領域の遺伝子型が表示される。RdRp領域、VP1領域のどちらか片方、もしくは両方にN.A.が表示された場合、新規遺伝子型の可能性があるので、できる限りの配列を決定し(RdRp領域はできるだけRdRp全長以上、VP1領域以降は、少なくともORF2全長のゲノム塩基配列)、論文をまとめるとともに、NoV S.C.に連絡を取り、新規遺伝子型番号をもらうと良い。
ORF1の部分配列、ORF2、3の部分配列の解析にもNoV遺伝子型分類ツールは対応しているので、ゴールデンスタンダードとして使用されているSKシリーズプライマー増幅産物を用いた場合でも、利用可能である。ORF1-2ジャンクション領域を含む200-250塩基程度の短い配列の場合、正確な遺伝子型分類が実行できない場合がある。その際には、配列長を延長してから再解析することを勧める。なお、NoV遺伝子型分類ツールでは、レポートのボタンをプッシュすることで分子系統樹を出力可能だが、この系統樹はML法や、ベイジアン法では無く、NJ法で描かれた系統樹なので注意が必要である。
d)NoVの命名法
“Proposal for a unified norovirus nomenclature and genotyping”で、推奨される標準株の遺伝子型表記方法は、ウイルス グループ/ホスト/分離国/分離年/ RdRp領域タイプ-VP1領域タイプ/株名、のように表記するのが望ましい。下記に表記例を示した。
“norovirus GII/Hu/FR/2004/GII.P12-GII.3/Paris23”
“norovirus GII/Hu/FR/2004/GII.12/Paris”
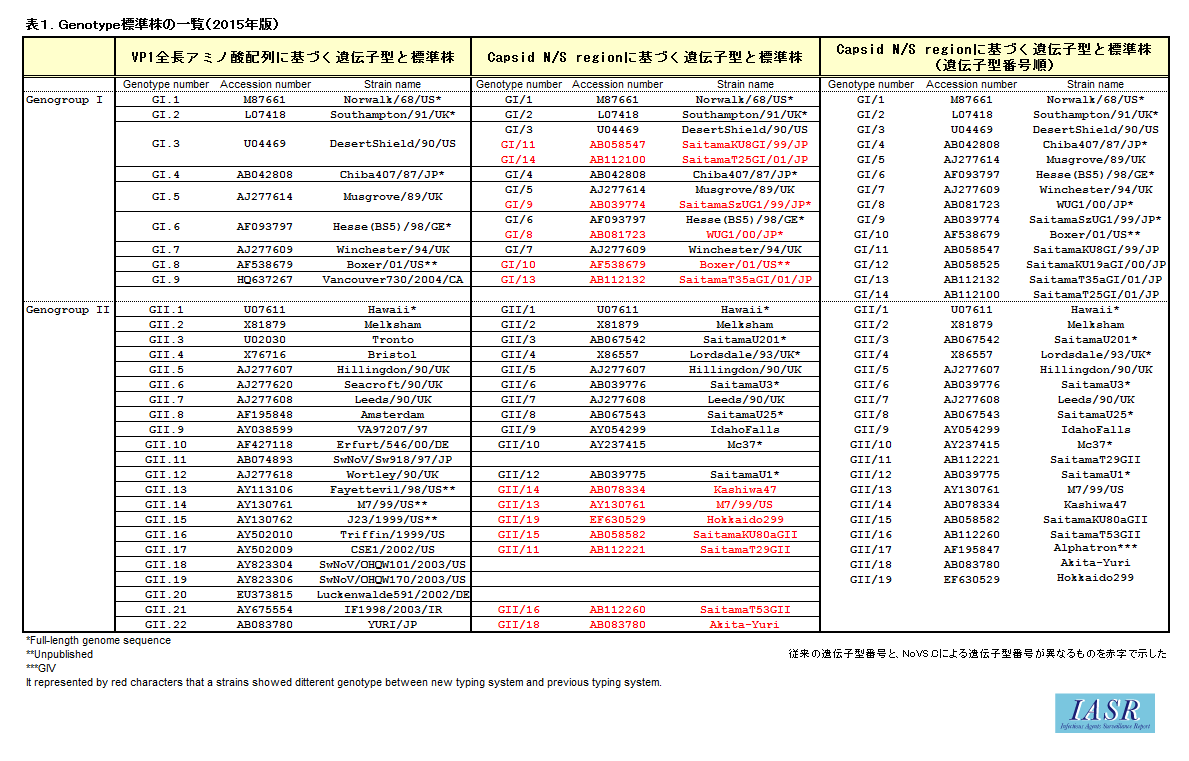
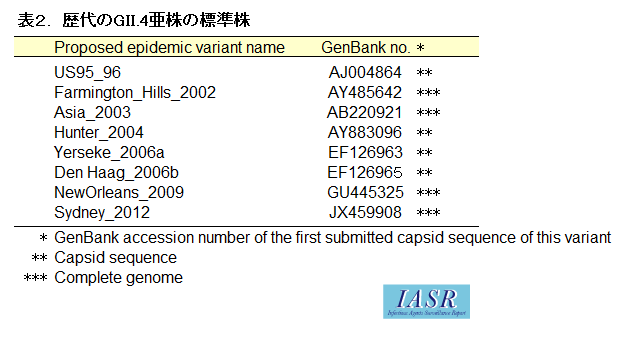
従来、国内ならびにIASRで用いられてきた表記方法と標準株、NoroNetによって提供されるNoV遺伝子型分類ツールによる標準株の関係を表1にまとめた。本改訂版では、特にGII.17に対応した。また、GII.4については、それぞれ歴代の亜株の標準株を表2にまとめた。
{kind=link}
{kind=link}
おわりに
NoroNetによるNoVの遺伝子型分類法は、本稿読者の研究者が論文をまとめる際に重要である。このガイドラインに従って株表記行うこと、表1、2に示したNoroNetの標準株を加えて分子系統解析を行うことが、国際的なNoVの分子系統解析のデータ共有を進めることに直結する。2015/16シーズンより、国内の厚生労働省の食中毒統計、感染症サーベイランス;NESID上の遺伝子型表記、IDWR, IASRで用いられてきたスラッシュ区切りの遺伝子型表記を一斉にNoroNetによるNoVの遺伝子型分類法の表記に切り替えていく(比較表参照)。研究者の皆さまには、ご迷惑をおかけして申し訳ないのだが、この切替作業への協力をお願いしたい。
参考文献
- Matsushima, et al., Euro Surveill, 20(26): pii=21173, 2015
- Khamrin P, et al., Euro Surveill, 20(28): pii=21185, 2015
- Katayama K, et al. (2002), Phylogenetic Analysis of the Complete Genome of 18 Norwalk-like Viruses, Virology 299(2): 225
- Kageyama T, et al. (2003), Broadly reactive and highly sensitive assay for Norwalk-like viruses based on real-time quantitative reverse transcription-PCR, J Clin Microbiol 41(4): 1548-1557
- Karst SM, Wobus CE, Lay M, Davidson J, & Virgin HWt (2003), STAT1-dependent innate immunity to a Norwalk-like virus, Science 299(5612): 1575-1578
- Kroneman A, et al. (2013), Proposal for a unified norovirus nomenclature and genotyping, Arch Virol
- Kroneman A, et al. (2011), An automated genotyping tool for enteroviruses and noroviruses, J Clin Virol 51(2): 121-125
- Sharp TM, Guix S, Katayama K, Crawford SE, & Estes MK (2010), Inhibition of Cellular Protein Secretion by Norwalk Virus Nonstructural Protein p22 Requires a Mimic of an Endoplasmic Reticulum Export Signal, PLoS ONE 5(10): e13130
- Eden JS, Sharpe LJ, White PA, & Brown AJ (2011), Norovirus RNA-dependent RNA polymerase is phosphorylated by an important survival kinase, Akt J Virol 85(20): 10894-10898
国立感染症研究所ウイルス第二部 片山和彦
同 感染症疫学センター 木村博一