ヒトノロウイルス(HuNoV)キャプシド遺伝子の分子進化について
(IASR Vol. 38 p.12-14: 2017年1月号)
1.はじめに
ノロウイルス(NoV)はカリシウイルス科ノロウイルス属に属するウイルスである。NoVのゲノムは, 3つのORF(open reading frame) を有し, 3つの構造蛋白と7つの非構造蛋白をコードしている。NoVはさらに, 主要抗原であるキャプシド蛋白をコードするキャプシド遺伝子の系統解析により7つの遺伝子群(GI-GVII)に分類され, ヒトでは主にGIおよびGIIが感染し, 急性胃腸炎を起こす。VP1遺伝子は, NoVのキャプシドをコードし, ウイルスの感染性や抗原性に関与する重要な蛋白である。本稿では, これまでの我々の既報1,2)を基に, ヒトNoV (HuNoV) GIおよびGIIのキャプシド遺伝子の分子進化について概説する。
2.ヒトNoV GIキャプシド遺伝子の分子進化
HuNoV GIキャプシド遺伝子の分子進化を明らかにするため, GenBankから網羅的にGIの各遺伝子型(GI.1~GI.9)のキャプシド遺伝子全長配列を入手した。さらに, 他のNoV種との共通起源に関するデータを得るため, ウシNoV GIII(BoNoV GIII)およびブタNoV GII(SwNoV GII)のキャプシド遺伝子全長配列を加え, 計69株(GI:65株, GIII:1株, GII:3株)をデータベースとし, ベイジアン・マルコフ鎖モンテカルロ法(MCMC法)を用いた時系列系統解析を行った。その結果, HuNoV GIとBoNoV GIIIの共通始祖ウイルスとSwNoV GIIの始祖ウイルスは約4,000年前に分岐したことが推定された(図1)。その後, 約3,000年前にHuNoV GIとBoNoV GIIIの共通始祖は分岐し, その後HuNoV GIはおおよそ800年の間に明瞭な2つの系統(lineage)および9つの遺伝子型に分岐したと推定された(図1)。また, 解析遺伝子の進化速度は, 1.25×10-3 substitutions/site/yearと非常に速いことがわかった。次に, 我々は既報と同様に複数の方法 (SLAC, FEL, IFELおよびMEME) を用いて正の選択(positive selection)・負の選択(negative selection)解析も行った3)。その結果, HuNoV GIキャプシド遺伝子では, 負の選択部位が400以上推定されるのに対し, 正の選択部位は一つもなかった。さらに, 時系列ゲノム動態の推移を解析する目的で, ベイジアン・スカイラインプロット (BSP) 解析を行った。BSP解析結果から, HuNoV GIでは約500年前から, 103以上のポピュレーションサイズを保ち続けていることから, このウイルスは高度にヒトに順化(adaptation)していることも推定された。
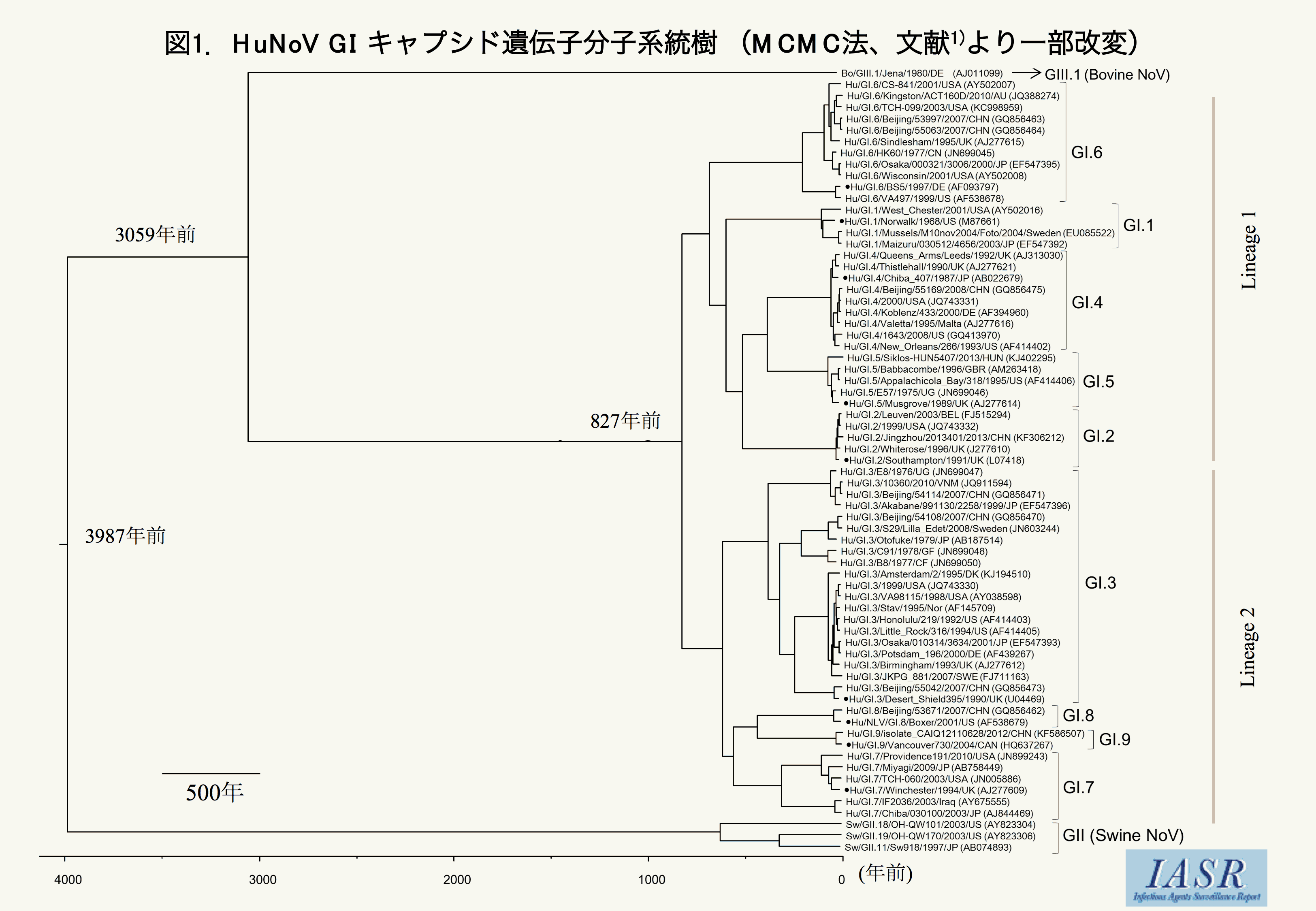{kind=link}
3.HuNoV GIIキャプシド遺伝子の分子進化
次に, HuNoV GIIキャプシド遺伝子の分子進化を明らかにするために, GenBankからHuNoV GIIの各遺伝子型(GII.1~GII.22)のキャプシド遺伝子を網羅的に入手した後, HuNoV GI, HuNoV GIV, SwNoV GIIおよびBoNoV GIIIを加えて, HuNoV GIと同様の解析を行った。図2に示すように, HuNoV GIとBoNoV GIIIの共通始祖ウイルスとSwNoV GIIおよびHuNoV GIVの共通始祖ウイルスが分岐した後, HuNoV GIIは約550年前にHuNoV GIVから分岐したことが推定された。その後さらにHuNoV GIIは, 約380年かけて, 3つのlineageおよび22の遺伝子型に分岐したと推定された。また, その進化速度は3.76×10-3 substitutions/site/yearとHuNoV GIと同程度の速度であることが推定された。また, 正・負の選択解析では, HuNoV GIIキャプシド遺伝子は, HuNoV GIと同様に400以上の負の選択部位を持ち, 2カ所の正の選択部位(aa6およびaa435)を有することが推定された。さらに, BSP解析ではHuNoV GIIは約380年前から102以上のポピュレーションサイズを持ちながら, ヒトに順化していることが示唆された。
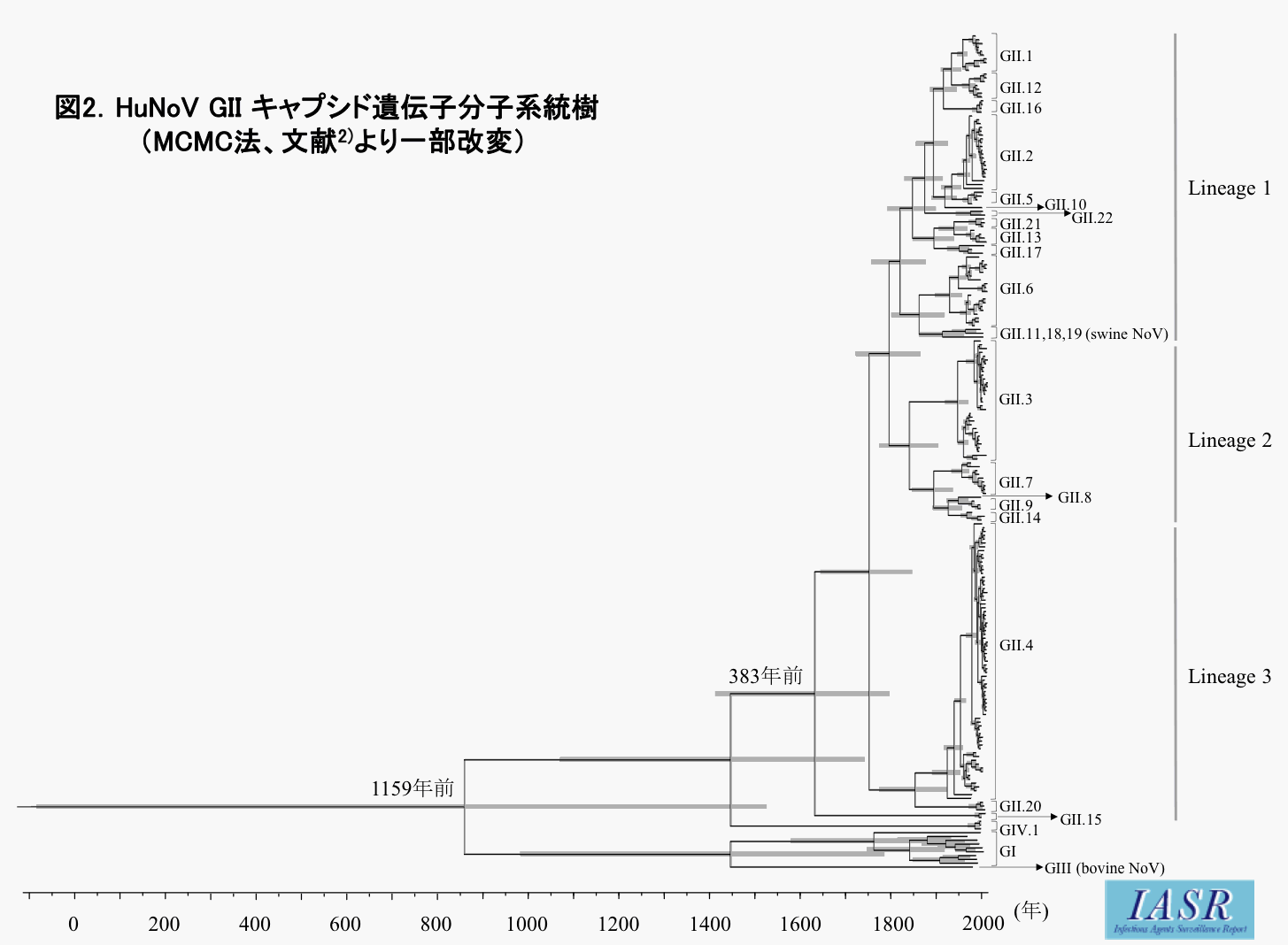{kind=link}
4.まとめ
これらの結果から, HuNoV GIとGIIは同じ胃腸炎の原因ウイルスではあるものの, それぞれ異なる種のBoNoV GIIIあるいはSwNoV GIIから分岐したと推定され, 長期間の時系列的進化を経て, ヒトの世界に順化し, 流行を広めてきたことが推定される。また, 正あるいは負の選択解析ではHuNoV GIキャプシド遺伝子には正の選択部位は認めなかったが, GIIでは2カ所有していた。この結果から, HuNoV GIIキャプシドはHuNoV GIに比し, より強い宿主免疫の選択圧を受けているとも考えられる。また, HuNoV GIとHuNoV GIIキャプシド遺伝子は, 遺伝学的多様性に富んでおり, 同じ遺伝子群でも遺伝子型により胃腸炎患者からの検出率に大きな差がある。今後, HuNoVの分子進化をより明らかにするためには, 遺伝子型別の詳細な解析を行い, 各遺伝子型におけるゲノム進化の特徴を詳細に解析していく必要があると考える。
参考文献
- Kobayashi M, et al., Sci Rep 5: 13806, 2015
- Kobayashi M, et al., Sci Rep 6: 29400, 2016
- Kimura H, et al., Infect Genet Evol 43: 398-406, 2016